

Mare Cyclicity
A solid understanding of mare cyclicity is the foundation on which to build or evaluate an equine breeding program. Horses differ from other species both in timing of cyclicity as well as endocrine patterns within a cycle. Basic principles can aid horse breeders in more effectively timing and breeding with or without hormone manipulation.
Seasonally Polyestrus
Mares are classified as seasonally polyestrous animals and are more generally termed “long day breeders.” This classification means mares cycle multiple times in the year but that these times are limited to when days are long. For the mare, this means conception prior to the hot days of summer and optimizing nutritional value for offspring by foaling early spring. This seasonality of the mare’s breeding cycle is mostly dictated by photoperiod; however, available food resources and temperature may also play minor roles.
Lighting signals, associated with the photoperiod, are interpreted by the pineal gland and ultimately converted into endocrine signals. At the center of this pathway is melatonin. Melatonin secretion increases during the night phase and quickly decreases at the beginning of the day phase. In seasonal breeders such as sheep (short day) and horses (long day), melatonin has a large impact on when cyclicity will end and ultimately resume. This pattern for long day breeders is evident in Figure 1.

In horses, low melatonin levels, which are associated with longer days, stimulate a cascade of hormones that ultimately control the ability of the mare to properly cycle. However, research indicates that estrous cycle control is not merely a result of melatonin exposure but also the pattern of its secretion. For this reason, a producer could use lighting regimes to artificially shorten days early in the year OR artificially lengthen days later in the year. Supplemental melatonin is also currently under investigation for artificial shortening of days early in the year.
Most horse breeders will opt to artificially lengthen days later in winter to induce an earlier start of cyclicity. If putting mares under artificial light, the perceived increase in day-length will not have mares cycling until 60–90 days later. To get as close to a January 1 birthday, but not before, most mares need to be under lights by December 15.
One important piece to understanding the seasonality of the mare is that she must undergo some period of shortened days in the year. This innate need for a period of anestrus (non-cycling) has been revealed in multiple research programs. In these programs, mares have been blinded or continually exposed to long days. Mares in both situations have responded by creating their own period of anestrus during the calendar year. For this reason, it is important for producers to monitor lighting on brood mares. Some irregularity in mare cyclicity has even been noted when mares are housed in heavily trafficked barns where artificial lighting stays on all or the majority of night hours.
Endocrine Control of the Estrous Cycle
Secretions modulating the estrous cycle are very different than those affecting seasonal cyclicity. It is necessary to understand what hormones act in the estrous cycle and the pattern in which they are secreted before developing a breeding program. First, the equine estrous cycle is separated into two distinct phases. These are the “luteal” phase and the “follicular” phase.
The luteal phase is dominated by the corpus luteum (CL) and the hormone progesterone, while the dominant follicle and the hormone estrogen control the follicular phase. The luteal phase controls the largest portion of the cycle, with approximately 14 days dedicated to the CL. The remaining 7 days of the 21-day estrous cycle make up the follicular phase. The time surrounding ovulation when a mare is in heat is a general guide that the mare is in the follicular phase. The time that a mare is not receptive to a stallion is the time frame when the luteal phase is dominating. Thereby, a producer can typically identify the phase a mare is in simply by teasing with a stallion.
The follicle is the structure that holds and is responsible for ovulating the female gamete or oocyte (egg). The secretion of estrogen from the follicle is in an effort to be more receptive to the male as well as to stimulate the cascade of events that ultimately result in ovulation. The CL is formed from remaining follicular tissue following its rupture at ovulation. During the transformation from follicular tissue to luteal, there is little hormone secretion from the gonads. Instead, the time period following ovulation is under control of residual estrogen. Once the transformation has finished, progesterone from the newly formed CL is intended to maintain pregnancy. This responsibility stays with the CL until no pregnancy is recognized or until approximately 120–150 days of gestation.
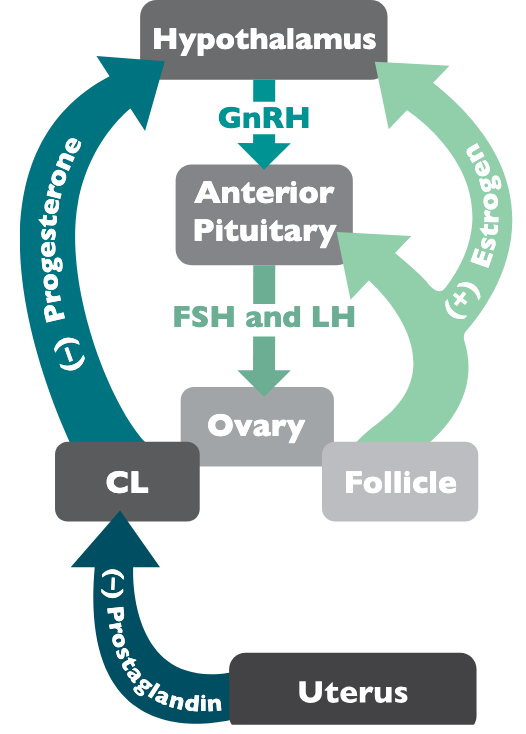
Cyclicity in the mare is a balance between and coordination among the hormones of the hypothalamus, pituitary gland, ovaries, uterine endometrium, and the pineal gland. How they communicate with each other ultimately determines the steps in the cycle. This communication can be either negative or positive in form, meaning that they can inhibit or promote hormone production from each other.
Gonadotropin releasing hormone (GnRH) is produced by the hypothalamus and acts on the anterior pituitary to stimulate the release of the follicle stimulating hormone (FSH) and luteinizing hormone (LH). The production of FSH is directly correlated with follicular recruitment and selection as well as the increase in circulating estrogen and increased follicular size. The production of LH is generally limited to increased secretion at the end of the cycle with its primary responsibility to stimulate exponential growth, maturation, and ovulation of the dominant follicle. The pattern of GnRH secretion determines the ratio of FSH to LH production throughout the cycle.
The pattern of GnRH production is ultimately determined by progesterone and estrogen. Progesterone feeds back to inhibit GnRH production, which skews the ratio towards FSH production (follicle recruitment).
Progesterone dominates approximately two-thirds of the mare cycle. The progesterone block is only removed with prostaglandin secretion from the uterine endometrium when an embryo is not recognized.
Prostaglandin regresses the CL and removes progesterone in circulation. When progesterone levels are diminished, the follicle is able to grow and increase estrogen production. Estrogen is different from progesterone in that it promotes GnRH production in a manner that is favorable towards LH production. The rise in LH causes the follicle to mature and then ovulate. These hormones all work in concert with one another to fulfill one complete mare cycle. The method by which these hormones and their production are controlled is briefly outlined in Figure 2.
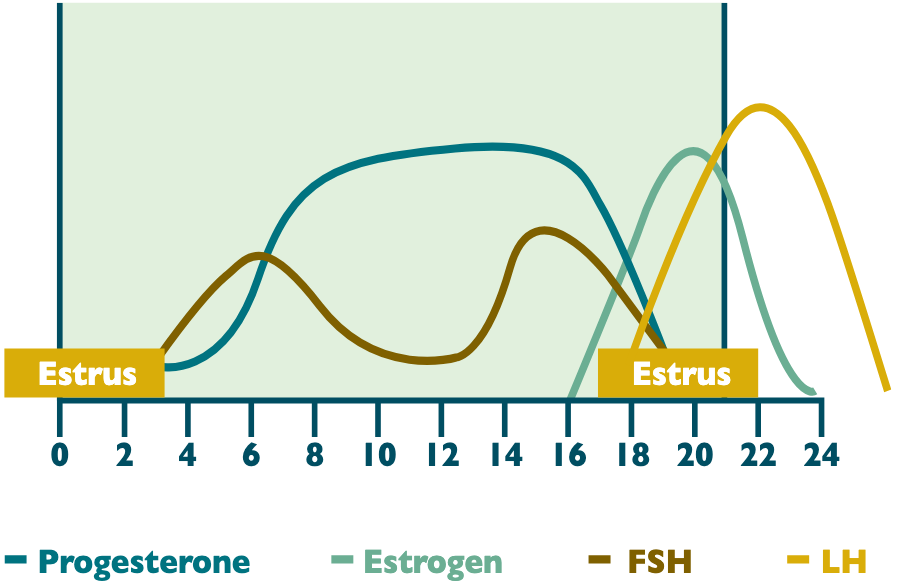
The estrous cycle in the mare is approximately 21 days but 3 days longer or shorter. The difference in estrous cycle lengths varies between mares, whereas each individual mare is relatively consistent in the length of time from one ovulation to the next.
Following ovulation is the transition from the follicular phase to the luteal. Progesterone concentrations begin to rise shortly following ovulation, but do not reach peak values until approximately 4 to 5 days post ovulation. During this time of progesterone rise, the CL is considered non-responsive to prostaglandin. Unique to the mare, LH remains elevated 1 to 2 days following ovulation. This longer duration of elevated LH is thought to be inherently important in CL development and function.
Generally, concurrent with the rise in progesterone post ovulation is the first rise in FSH. This first follicular recruitment does not result in ovulation because of its coordinated time with an active progesterone block. Progesterone remains elevated for 14–15 days. If no conceptus is recognized, the progesterone block is not maintained as prostaglandin from the uterine endometrium causes regression of the CL. This removal of the progesterone block comes just after a second, more pronounced follicular wave that occurs approximately 10 days before ovulation.
With circulating progesterone minimizing, estrogen is allowed to build in the dominant follicle. Concurrent reduction in progesterone and a rise in estrogen lends to higher pulse frequency of GnRH and the resulting preferential production of LH. This increase in LH then acts in a positive feedback loop that is self-amplifying. Self-amplification simply means that LH feeds itself in order to increase its own production. In this self-amplification loop, estrogen promotes LH production. This rise in LH then promotes increased follicular growth, and ultimately that follicular growth increases estrogen secretion, which again promotes LH production. A decline in progesterone and marked increase in estrogen, as a result of follicular growth, also indicates the beginning of estrus. Estrogen is directly responsible for the secondary sex characteristics that are observed in a mare when she is in estrus or heat. This time is when she is physically receptive to the stallion as most breeders find when teasing. Certain situations such as having a foal at her side or improper body conditioning, may cause the heat to be silent or without secondary sex characteristics.
Normally estrus lasts 5 to 7 days but can be as short as 2 or as long as 12 days in length. At times a mare may also exhibit signs of estrus but will be unable to ovulate. These are most commonly found in mares during the early to mid-transitional period. During the early spring and late summer, longer periods of estrus are seen in mares transitioning in and out of season. The shortest estrus occurs in mares during the summer solstice (the longest day of the year). These markers are important for breeders using teasing as a sole identifier of mares in heat.
If using the average of 7 days for the length of time mares are in estrus, then there are approximately 5 days from the onset of estrus until ovulation. The surge in LH occurs approximately 3 days after estrus begins and, thus, is 2 days prior to ovulation. Mares may continue to tease in 1 to 2 days following ovulation as a result of low progesterone and residual estrogen effects from the ovulated dominant follicle. The general pattern of hormone release during the estrous cycle of the mare can be found in Figure 3.
Follicular Dynamics in the Mare
Follicular Development
Folliculogenesis, the process by which immature follicles develop into pre-ovulatory follicles, has four distinguishable follicular groups. The differentiation between follicular phases is based on changes in basic characteristics to include size and cellular layers. The first distinct follicular group is primordial follicles that are immature in their developmental status. Primordial follicles, with a single layer of squamous cells, develop into primary cells, which have a single layer of cuboidal cells also referred to as follicular cells. From this point, the follicle has the ability to continue on the developmental pathway to a secondary follicle or degenerate with removal from the follicular pool in a process called atresia.
If development continues, there will be the addition of one or more follicular cell layers. The next generational development of the follicle is the tertiary follicle, which is no longer classified as microscopic. The increase in size attributed to this developmental stage is the result of an “antrum” or fluid filled cavity. Hence, you may also hear references to these follicles as the antral follicles. The antrum, or fluid filled cavity, is the origination point for the hormone estrogen following selection of these follicles.
The antral follicles in the mare may be classified as small (less than 10 mm), medium (10–20 mm), and large (greater than 20 mm) to help reproductive personnel determine their current status. If the mare’s typical follicular size at ovulation is known, some of these numbers for size classification may be skewed to fit the mare. The fate of the tertiary or antral follicles is the dynamics of the follicular phase of the estrous cycle in the mare. These follicles will go through three phases of recruitment and selection from which typically only one will be selected for overall dominance. The follicles not in that final selection for dominance will degenerate.
Follicular Waves
Following follicular recruitment, antral follicles have endocrine capacity and the ability to secrete estrogen. This reproductive event is often termed a “follicular wave” and may or may not result in ovulation. Estrogen from recruited antral follicles act in a positive feedback mechanism to the HPG axis to stimulate additional production of FSH and LH, which “feed” the recruited follicles further. Of these recruited follicles, few are then selected to undergo further growth. This pattern of growth and degeneration are the symbolic “wave.”
Horses are known for their variation in types of follicular waves. The three types of waves are: minor wave, major anovulatory wave, and major ovulatory wave. Figure 4 depicts possible waves found in horses as reported by a research team from Israel.

A minor wave is characterized by the inability for a follicle to achieve dominance, whereas a major wave has both a dominant follicle and subordinate follicles. Typically, horses have one to two waves per cycle, and wave types can be influenced by pregnancy, stage of the estrous cycle, season, breed, and age. The culmination of a major ovulatory follicular wave is the ovulation of typically one follicle.
Wave patterns are highly variable across breeds and within animals. Typically, Thoroughbreds are known for two follicular waves, while Quarter Horses and ponies are noted for having only a singular follicular wave. Determining follicular wave patterns can be achieved through follicular mapping via ultrasound.
Follicular Dominance and Mapping
If conducting periodic ultrasound scans, mapping follicular development and wave patterns can help a producer understand where a mare is currently in her estrous cycle. The ultimately goal in follicular mapping would be to identify and track the dominant follicle. The dominant follicle is the follicle that will later ovulate. The process by which a follicle is capable of achieving dominance and understanding the conditions surrounding this dominance will allow a breeder to more accurately concentrate breeding activities.
It is assumed following first identification of small, antral follicles, that they will grow at similar rates for approximately 6 days. From this smaller group of follicles, there are two follicles, generally in the mare, which will continue to grow larger as the cycle progresses. Eventually, one of these follicles will become dominant. This point in time is referred to as deviation. At deviation, typically, one follicle goes on to grow larger while the other become subordinate (Figure 5).
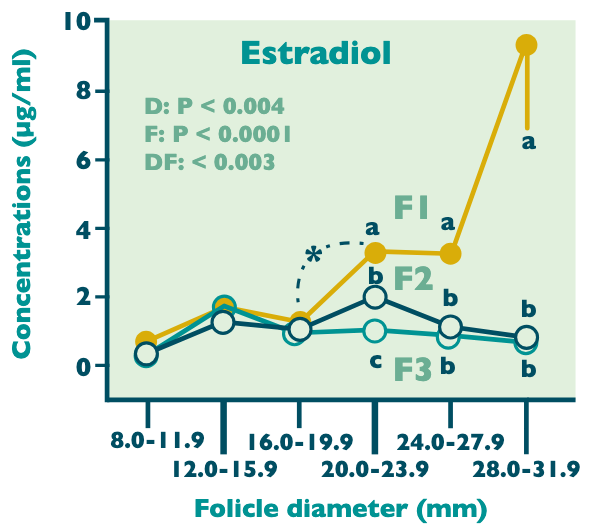
The time from peak FSH in the cycle to deviation is approximately 3 days. How a singular follicle achieves dominance is not well understood; however, it is believed that the dominant follicle is more responsive to low levels of circulating FSH. These follicles that are capable of responding to lower FSH appear to have an increased estradiol concentration prior to deviation.
This period in time surrounding follicular recruitment, selection, and dominance is marked by alterations in various hormone levels. The emergence of a follicular wave is marked by a surge in FSH. The secretion of FSH remains elevated until a point in time when the largest follicle reaches a size of 13–15 mm and begins secreting inhibin. Inhibin is produced by the granulosa cells of the developing follicle and acts directly on the anterior pituitary to inhibit FSH production. This suppression of FSH is solely assumed by the dominant follicle around the time of deviation through a rise in both estrogen and inhibin.
Concurrent with the dominant follicle’s suppressive action is its acquisition of LH receptors within the granulosa cells. With the low circulating FSH in an effort to make all other follicles subordinate, the dominant follicle has responded accordingly to depend on LH for final growth and maturation. Recent research suggests that the timeframe in which deviation occurs is when the two largest follicles reach approximately 20–24 mm in diameter. At this point, one follicle will continue to grow while the other(s) will see no further growth. This follicle then becomes the dominant, pre-ovulatory follicle.
Understanding the phenomenon and dynamics of follicular waves will help horse breeders further understand the changes seen externally. The ability to directly locate and track the growth of specific follicles will assist a breeder in understanding cycle stage and breeding potential. Knowledge of follicular wave patterns, hormones of the cycle, and the potential for exogenous hormone manipulation may enable one to breed more effectively and accurately.
The mapping and growth of the dominant follicle will serve the greatest purpose to most producers as ultrasounding will not commence in a breeding scheme, generally speaking, until a mare is teasing in. The growth of the dominant follicle can range from 2–5 mm per day. Uniquely, this growth ceases some 24–48 hours prior to ovulation. During this same timeframe, diagnostic examinations with ultrasound can pinpoint a change in follicular shape. The follicle, prior to ovulation will transition from a spherical shape to one that might have a point or cone-like side.
This tracking and mapping may reduce the number of times a mare is bred as ovulation will generally occur prior to the cessation of estrus (0–48 hours). Oocyte viability post ovulation is approximately 6 hours, while sperm viability is closer to 48 (fresh or cooled semen). Therefore, inseminating post ovulation is often a wasted effort. If using frozen semen, the duration of viability for semen is greatly reduced (approximately 12 hours) and a mare may require more than one ultrasound imaging per day.
Typically, producers will choose to start thinking about breeding once follicle size has reached 35 mm. However, producers must be aware that there is large variability in follicular size at ovulation based on the breed, total number of follicles, and how many previous cycles the mare has had in the current breeding season. In general, draft breeds naturally ovulate larger follicles. If there are two pre-ovulatory follicles, follicular size at ovulation may be 3–5 mm smaller. Multiple follicles may ovulate at a size less than 25 mm as well. Additionally, mares are more likely to ovulate a larger follicle (by 4–8 mm) early in the breeding season (spring) with smaller follicles ovulated later in the season (summer). The variability in follicular size based on season follows the trend for longer periods of estrus early in the season. To reduce variability and further define the timing of estrus and ovulation, exogenous hormone administration may be employed.
Use of Exogenous Hormonal Manipulation
There are several hormones available for exogenous administration in the mare. With an understanding of the mare estrous cycle and follicular dynamics, timing of hormone administration will become more logical. Exogenous hormones that may prove useful in an equine breeding program include: progesterone, prostaglandin, human chorionic gonadotropin (hCG), GnRH, and FSH. The latter hormone is used for superovulation programs for embryo transfer and will not be discussed for the traditional brood mare.
Progesterone
Inhibits the rise in LH and thus inhibits heat/teasing activity. The administration of progesterone serves to mimic the presence of a CL and may be administered with or without the presence of a CL. Progesterone supplementation allows for the pooling or build-up of LH for ovulation induction with supplementation lengths varying from 10–15 days. This pooling allows for progesterone administration to both ease the transition of mares into the breeding season as well as more effectively determine when a mare will be in estrus. With its administration and subsequent removal, ovulation generally follows given a follicle of sufficient size (greater than or equal to 25 mm).
When administered to cycling mares, this program should include a prostaglandin injection at the conclusion of progesterone treatment. This will remove the potential for natural progesterone production by a previously developed CL. Mares that are under the direction of this program (progesterone plus prostaglandin) are generally in estrus within 2 to 4 days. Realize that this more effectively times when mares will be in heat but does not control timing of ovulation.
Progesterone may be administered at any point during the estrous cycle as evident in Figure 6. Progesterone is often given to performance mares to reduce behavioral and some physical issues associated with being in heat.

Prostaglandin
Responsible for the regression of the CL and removal of circulating progesterone, prostaglandin is only effective at inducing a response when a CL is present. An active CL is generally present on approximately days 6 to 18 of the cycle (Figure 7).

Administration of this hormone will shorten the period of luteal control and result in a faster return to estrus—often termed “short cycling.” A producer can expect a mare to return to estrus or heat, on average, 4 to 5 days following injection. When combined with preceding progesterone treatment, as discussed above, a mare can be expected to return to heat slightly early at approximately 2 to 4 days following injection.
An important fact to remember is that early pregnancy is dependent on progesterone for survival—administration of prostaglandin to a pregnant animal will result in embryonic loss. Only administer to animals not previously bred or classified as not pregnant. Prostaglandin will not be effective in animals during estrus or just following ovulation. An active CL is necessary for proper response and may be identified through teasing or ultrasonography. Responsiveness to prostaglandin will be evident through extensive sweating and potential abdominal discomfort. Some products advertise for a reduction in these side effects.
Human Chorionic Gonadotropin (hCG)
Extremely efficient at inducing ovulation because of its long half-life. Most mares will need to have a 35 mm follicle or greater to respond to hCG. Resulting ovulation is likely to occur a few hours sooner following hCG administration when compared with GnRH. Time from administration to ovulation is approximately 36–42 hours.
This earlier response is likely a result of direct action by hCG on the ovary. In addition to a faster response, this direct action appears to be more efficient than GnRH when used on mares early in the season. Later in the seasons, mares are likely to respond faster, possibly in as little as 24 hours, to hCG administration.
The main concern with hCG is related to diminished effectiveness as the season progresses. The reality is that the foreign nature of this substance (human) in the mare tends to result in the building of antibodies. This buildup may decrease its effectiveness if repeatedly used in a breeding season.
GnRH
Can be used more effectively on a smaller follicle (30 mm) if the breeder desires to speed up the estrous cycle. The window from administration until ovulation can be longer than with hCG, averaging approximately 44–48 hours. The longer duration from hormone administration until ovulation is the result of an additional step required, since GnRH must act through the pituitary to stimulate hormone release and take effect.
Timing of administration for both hCG and GnRH is more limited than for progesterone and prostaglandin (Figure 8).

If most look at why these hormones are utilized, it is to answer the “unknowns.” Progesterone can help answer the question of “is she going to come into heat soon?” Progesterone and prostaglandin can help answer the question of “when is she going to come into heat next?” GnRH and hCG can help answer the question of “when is she going to ovulate?” The main feature is that they can more accurately tell you “when will I be breeding her again?”
As a final thought on hormone manipulation, realize that these hormones may be used as individuals to manipulate certain points within a cycle or together to more precisely sequence the cycle.
The Big Picture
For current or potential horse breeders, understanding the physiology of the mare will help you to become more comfortable with breeding events and confident in the choices you make regarding them. A sound understanding of the reproductive endocrinology of your mare will allow you to become a more effective breeder with or without the use of hormone manipulation. The final item to remember is that not all mares read the same book so expect differences between them in both behavior and responses.






