

Introduction
Calcium (Ca2+) is an essential macronutrient involved in key physiological and developmental processes in plants, including cell development, membrane permeability, and cell wall structure. Ca2+ has low mobility and primarily moves through the xylem via the transpiration stream.
The transport of Ca2+ to plant organs depends on the transpiration rate of the organ. Leaves generally have a higher transpiration rate (the rate at which water is released into the atmosphere) than fruit, because leaves have a high number of stomata (pores). Therefore, Ca2+ accumulation in leaf tissues is higher than in fruit tissues.
Did you know?
Stomata are microscopic pores on leaves that allow plants to regulate water loss and gas exchange. The stomata located in the berry lose functionality as the berry develops, allowing less transpiration to occur in the berry. This leads to less Ca2+ accumulation in the berry.
Environmental factors (e.g., temperature, light, moisture) also influence Ca2+ uptake and transport within the plant. Higher temperatures and light intensity may increase transpiration rates, thereby facilitating the movement of Ca2+ toward the leaves. However, extremely high temperatures or insufficient water availability may affect Ca2+ movement.
Low relative humidity increases transpiration in leaves, and high humidity restricts it, affecting Ca2+ translocation to leaves and fruit. Additionally, drought conditions can limit Ca2+ translocation to the fruit.
Related Content
Blueberry plants grow in soil with low pH, typically within a range of 4.0 to 5.5. Under such conditions, even if soil Ca2+ levels are sufficient, Ca2+ may be displaced by other cations (positively charged atoms or molecules)—such as hydrogen (H+), aluminum (Al3+), and manganese (Mn2+)—limiting its availability for plant uptake.
Issues With Calcium Deficiency in Blueberries
Although blueberries have a low calcium requirement, blueberry plants can exhibit Ca2+ deficiency symptoms, including necrotic spotting and interveinal chlorosis in young leaves. Calcium deficiency in the berry is not commonly reported, but a physiological disorder called “green fruit drop” has been related to low Ca2+ levels in the berries and high nitrogen (N) fertilization rates. Green fruit drop disorder causes lower fruit production, and as the berries ripen, internal browning develops, although it is not visible on the exterior of the fruit. Green fruit drop is commonly reported in ‘Draper’, a northern highbush blueberry (NHB) grown in colder regions.
It is common for blueberry growers to perform foliar calcium applications a week or two before harvest. However, the effectiveness of these applications is linked to factors such as the application rate, timing, the growth stage of the plant, and the product applied.
In northern highbush blueberries, calcium accumulation stops or slows down at the late green fruit stage, and it is minimal during fruit ripening. Therefore, Ca2+ application after the early green stage might not increase Ca2+ content in the berry. However, there is a lack of information on fruit calcium concentration for rabbiteye and southern highbush blueberry (SHB) cultivars, the two types of blueberries grown in Georgia. This resource focuses on quantifying the Ca2+ concentrations at different stages of berry development and within the berry.
Calcium Concentration Study Methods
For this study, three different commercial fields planted with the SHB cultivar ‘Farthing’ and the rabbiteye ‘Brightwell’ and ‘Powderblue’ cultivars were sampled starting during full bloom (full bloom, S6, to fruit coloring, S10; see Figure 1) and analyzed for Ca2+ concentration.

At the full bloom stage (S6) and at the petal-fall stage (S7), the whole flower was sampled, including the ovary and surrounding floral tissues. From the early green stage (S8) to the fruit coloring stage (S10), only the developing fruit (ovary) tissue was used for Ca2+ quantification. In addition, leaf tissue and soil samples were collected and analyzed.
For Ca2+ berry concentration, 20 berries per cultivar were harvested at 25% blue stage. The pulp, peel, and seeds were separated for Ca2+ quantification in individual tissues, and the number of seeds per berry was also quantified for each cultivar.
Calcium Concentration at Different Growth Stages
Concentration of Ca2+ was higher during the early stages of fruit development in the ‘Farthing’, ‘Brightwell’, and ‘Powderblue’ cultivars. Across all cultivars, there was a decline in Ca2+ concentration after the petal-fall stage (S7; see Figure 2).
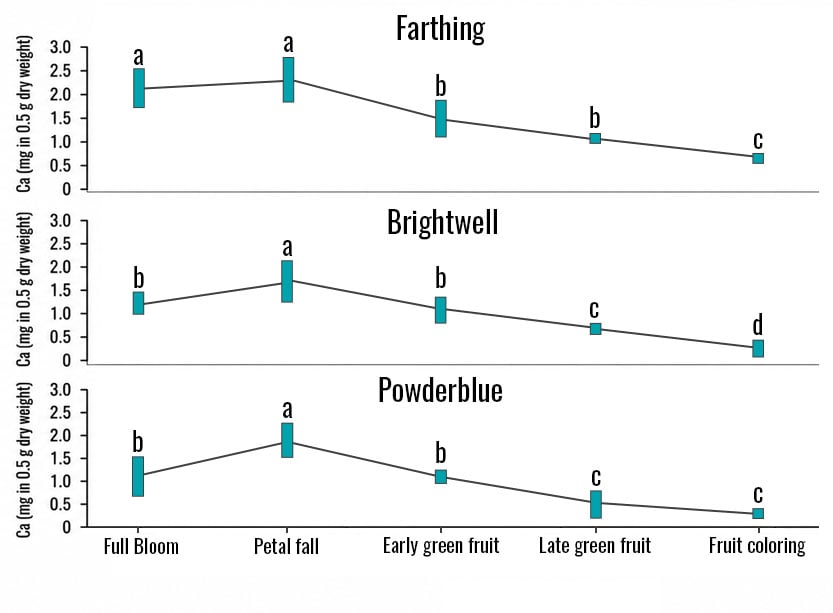
The decrease in Ca2+ concentration started at the early green fruit stage (S8) and may have resulted from the differences in transpiration rates between the leaves and the fruit. Leaves usually have higher transpiration rates, which allow for higher Ca2+ translocation and accumulation in this organ. Additionally, an increase in cuticle thickness at later stages of fruit development limits transpiration rate in the berry and consequently limits Ca2+ translocation to the fruit.
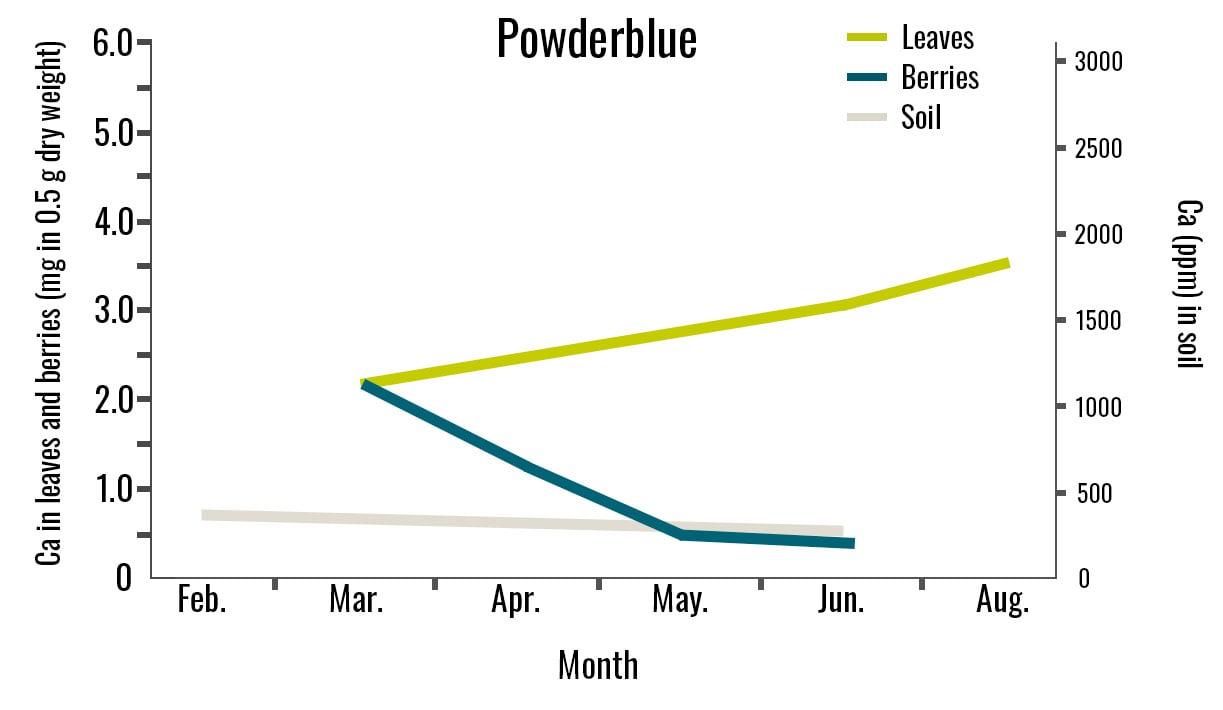
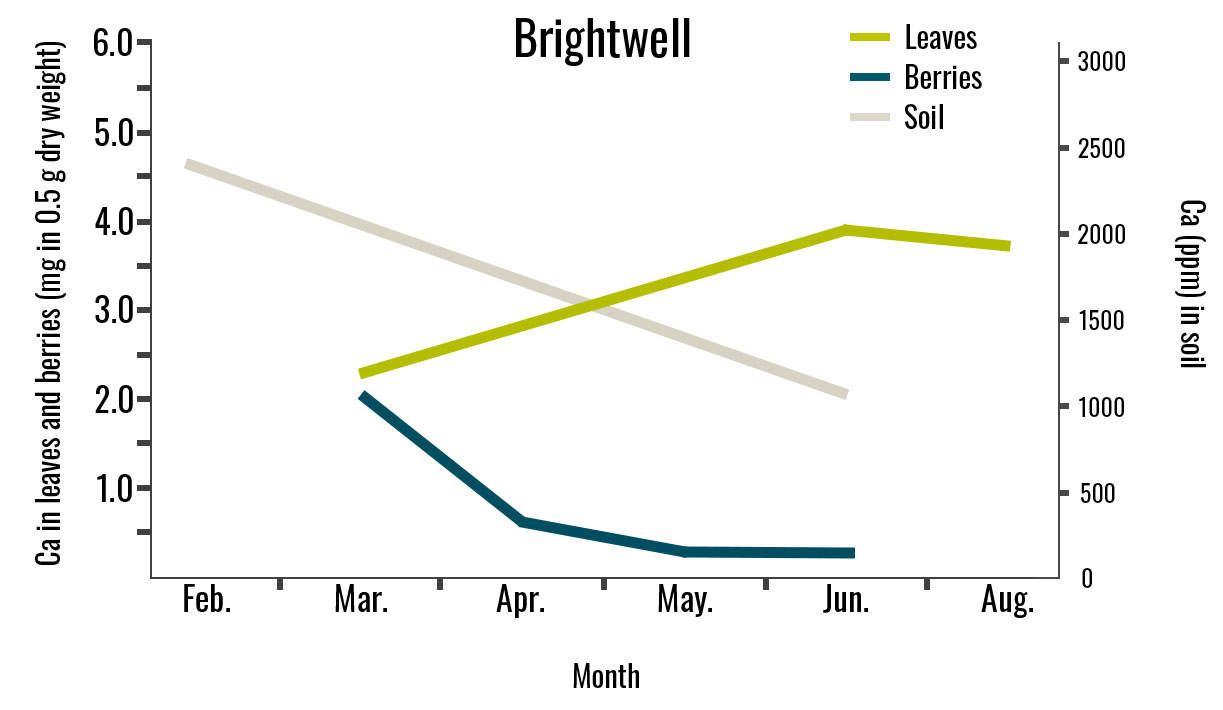
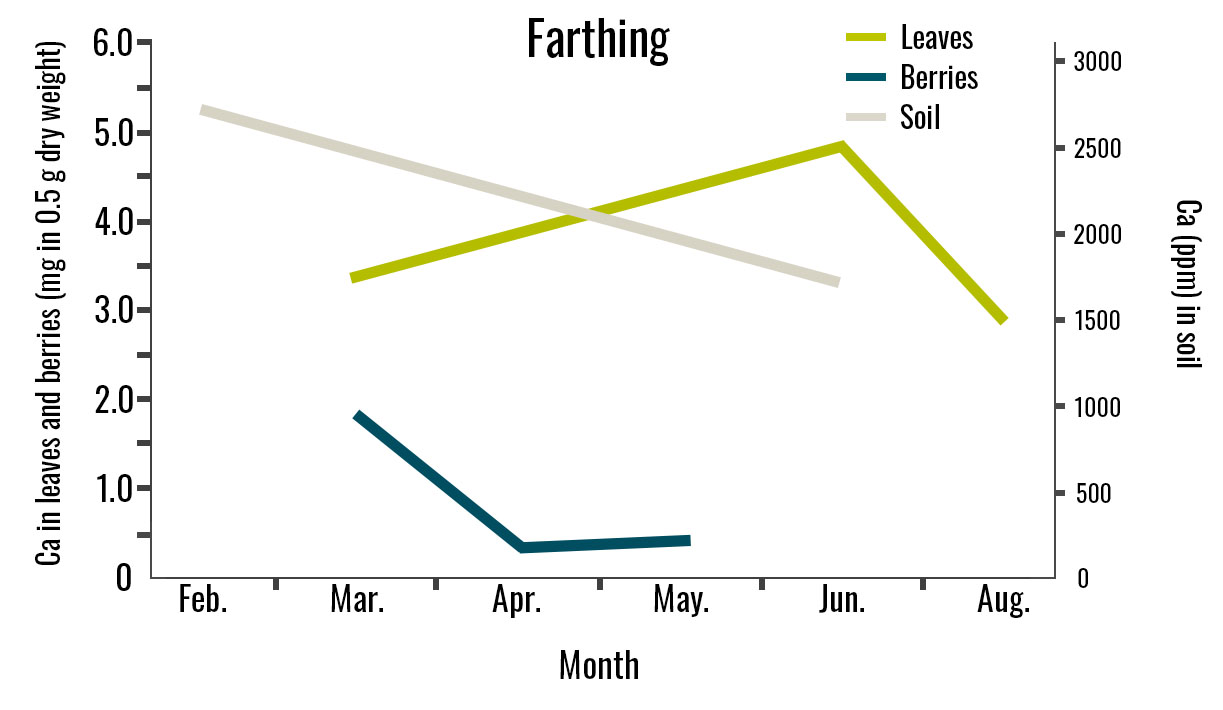
Figure 3. Calcium Concentration in Leaves, Berries, and Soil for Three Cultivars. Leaf samples were collected in March, June, and August. Berry samples were collected from March to June, and soil samples were collected in February and June. This study was conducted in 2024 in Alma, GA. Leaf and berry Ca2+ values are expressed as milligrams in 0.5 g of dry tissue. For each analysis, 0.5 g of dry tissue was used. Each replicate consisted of 40 mature leaves and 30 berries.
Calcium in the Leaves and Soil
The Ca2+ concentration in leaves and berries was similar across all three cultivars, although it varied over time for each cultivar. In ‘Farthing’, ‘Brightwell’, and ‘Powderblue’ berries, calcium concentration declined from March to April, while leaf calcium concentration increased from March to June (Figure 3). However, leaf Ca2+ concentration decreased in August for ‘Farthing’ and ‘Brightwell’ but increased for ‘Powderblue’ despite the low Ca2+ concentration in the soil.
Changes in transpiration rates in the plant may have influenced Ca2+ accumulation in the leaves. High transpiration rates allow plants to uptake calcium more efficiently, which may explain the higher leaf calcium concentration even when calcium in the soil was low.
In August, ‘Farthing’ presented a decrease in leaf Ca2+ concentration, which may be related to reduced transpiration under high-temperature conditions. The average high temperature in Alma, GA, during August ranges from 89.0 °F to 92.8 °F, which can cause stomata closure to prevent excessive water loss, slowing down leaf transpiration and limiting leaf Ca2+ uptake.
Among the cultivars, ‘Powderblue’ exhibited the lowest soil Ca2+ concentrations (Figure 3); the soil pH measured at this farm was more acidic (3.9–4.1) during both sampling months. In contrast, the soil pH for ‘Farthing’ ranged from 3.9 to 7.3, while ‘Brightwell’ ranged from 4.0 to 6.5, which might allow for higher Ca2+ in the soil solution.
Calcium Concentration in the Berry
For Ca2+ concentration within the berry, only the ‘Farthing’ and ‘Brightwell’ cultivars were evaluated. Ca2+ concentration in the pulp, peel, and seeds was higher in ‘Farthing’ compared to ‘Brightwell’. The tissue with the highest Ca2+ in ‘Farthing’ was the pulp, while in ‘Brightwell’ it was the seeds (Figure 4). In NHB cultivars, higher Ca2+ concentration in the peel and seeds has been reported, although these patterns can vary by cultivar.
Figure 4. Calcium Concentration in Tissues of Two Cultivars. Berries were harvested at the ripe stage in 2024 in Alma, GA. Peel, pulp, and seeds from 20 berries per cultivar were pooled for analysis. Values are expressed as milligrams in 0.5 g of dry tissue.
The difference in the number of seeds between cultivars was not statistically significant (Figure 5). However, ‘Brightwell’ has more seeds than ‘Farthing’. The differences in the number of seeds among the cultivars are linked to factors such as pollination performance and environmental conditions during pollination, which can influence Ca2+ concentration within the berries.
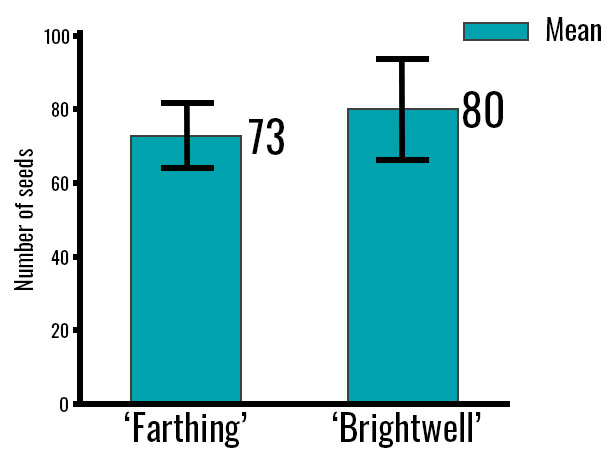
Thus, the greater Ca2+ concentration found in the seeds of ‘Brightwell’ could be associated with its higher seed number compared with ‘Farthing’. A larger number of seeds may increase sink strength, leading to a higher Ca2+ concentration in this tissue.
Conclusion
In the three cultivars studied, Ca2+ concentrations peaked during the petal-fall stage and subsequently began to decline as the berries ripened. Strategies aimed at increasing Ca2+ concentrations in the berries are likely to be most effective if applied early in the fruits’ developmental stages. Ca2+ concentration within the berries differs among cultivars and might also be affected by the environmental conditions during berry development and ripening.
References
Arrington, M., & DeVetter, L. W. (2017). Foliar applications of calcium and boron do not increase fruit set or yield in northern highbush blueberry (Vaccinium corymbosum). HortScience, 52(9), 1259–1264. https://doi.org/10.21273/HORTSCI12207-17
Arrington, M., & DeVetter, L. W. (2018). Increasing honey bee hive densities promotes pollination and yield components of highbush blueberry in western Washington. HortScience, 53(2), 191–194. https://doi.org/10.21273/HORTSCI12644-17
Bramlage, W. J., Weis, S. A., & Greene, D. W. (1990). Observations on the relationships among seed number, fruit calcium, and senescent breakdown in apples. HortScience, 25(3), 351–353. https://doi.org/10.21273/HORTSCI.25.3.351
Bryla, D. R., Orr, S. T., DeVetter, L. W., & Yang, W. Q. (2022). Practices for increasing calcium content and improving fruit quality and shelf life of blueberries. Acta Hortic., 1381, 309–316. https://doi.org/10.17660/ActaHortic.2023.1381.40
Buccheri, M., & Di Vaio, C. (2005). Relationship among seed number, quality, and calcium content in apple fruits. Journal of Plant Nutrition, 27(10), 1735–1746. https://doi.org/10.1081/PLN-200026409
Buckley, T. N. (2005). The control of stomata by water balance. New Phytologist, 168(2), 275–292. https://doi.org/10.1111/j.1469-8137.2005.01543.x
Choi, J. H., Chung, G. C., & Suh, S. R. (1997). Effect of night humidity on the vegetative growth and the mineral composition of tomato and strawberry plants. Scientia Horticulturae, 70(4), 293–299. https://doi.org/10.1016/S0304-4238(97)00055-1
Dogterom, M. H., Winston, M. L., & Mukai, A. (2000). Effect of pollen load size and source (self, outcross) on seed and fruit production in highbush blueberry cv. ‘Bluecrop’ (Vaccinium corymbosum; Ericaceae). American Journal of Botany, 87(11), 1584–1591. https://doi.org/10.2307/2656734
Doyle, J. W., Nambeesan, S. U., & Malladi, A. (2021). Physiology of nitrogen and calcium nutrition in blueberry (Vaccinium sp.). Agronomy, 11(4), 765. https://doi.org/10.3390/agronomy11040765
Ferguson, I. B. (1984). Calcium in plant senescence and fruit ripening. Plant, Cell & Environment, 7(6), 477–489. https://doi.org/10.1111/j.1365-3040.1984.tb01438.x
Gerbrandt, E. M., Mouritzen, C., & Sweeney, M. (2019). Foliar calcium corrects a deficiency causing green fruit drop in ‘Draper’ highbush blueberry (Vaccinium corymbosum L.). Agriculture, 9(3), 63. https://doi.org/10.3390/agriculture9030063
González-Villagra, J., Ávila, K., Gajardo, H. A., Bravo, L. A., Ribera-Fonseca, A., Jorquera-Fontena, E., Curaqueo, G., Roldán, C., Falquetto-Gomes, P., Nunes-Nesi, A., & Reyes-Díaz, M. M. (2024). Diurnal high temperatures affect the physiological performance and fruit quality of highbush blueberry (Vaccinium corymbosum L.) cv. Legacy. Plants, 13(13), 1846. https://doi.org/10.3390/plants13131846
Guo, S., Abudureheman, R., Zhang, Z., Zhong, H., Zhang, F., Wang, X., Nasir, M., & Wu, J. (2025). Physiological responses of grapevine leaves to high temperature at different senescence periods. Plants, 14(20), 3142. https://doi.org/10.3390/plants14203142
Inoue, K., Yokota, H., & Yamada, Y. (1988). Effect of Ca in the medium on root growth under low pH conditions. Soil Science and Plant Nutrition, 34(3), 359–374. https://doi.org/10.1080/00380768.1988.10415692
Hanson, E., & Hancock, J. F. (1996). Managing the nutrition of highbush blueberries. Michigan State University Extension.
Hawkesford, M. J., Cakmak, I., Coskun, D., De Kok, L. J., Lambers, H., Schjoerring, J. K., & White, P. J. (2023). Functions of macronutrients. In E. Rengel, I. Cakmak, & P. J. White (Eds.), Marschner’s Mineral Nutrition of Plants (4th ed., pp. 201–281). Academic Press. https://doi.org/10.1016/B978-0-12-819773-8.00019-8
Jing, T., Li, J., He, Y., Shankar, A., Saxena, A., Tiwari, A., Maturi, K. C., Solanki, M. K., Singh, V., Eissa, M. A., & Ding, Z. (2024). Role of calcium nutrition in plant physiology: Advances in research and insights into acidic soil conditions – A comprehensive review. Plant Physiology and Biochemistry, 210, 108602. https://doi.org/10.1016/j.plaphy.2024.108602
Kabir, M. Y., & Díaz-Pérez, J. C. (2025). Calcium route in the plant and blossom-end rot incidence. Horticulturae, 11(7), 807. https://doi.org/10.3390/horticulturae11070807
Karley, A. J., & White, P. J. (2009). Moving cationic minerals to edible tissues: potassium, magnesium, calcium. Current Opinion in Plant Biology, 12(3), 291–298. https://doi.org/10.1016/j.pbi.2009.04.013
Kirkby, E. A., & Pilbeam, D. J. (1984). Calcium as a plant nutrient. Plant, Cell & Environment, 7(6), 397–405. https://doi.org/10.1111/j.1365-3040.1984.tb01429.x
Lobos, T. E., Retamales, J. B., Luengo Escobar, A., & Hanson, E. J. (2021). Timing of foliar calcium sprays improves fruit firmness and antioxidants in “Liberty” blueberries. Journal of Soil Science and Plant Nutrition, 21, 426–436. https://doi.org/10.1007/s42729-020-00371-2
Marschner, P. (Ed.). (2012). Marschner’s mineral nutrition of higher plants (3rd ed.). Academic Press. https://doi.org/10.1016/C2009-0-63043-9
Münzbergová, Z., & Skuhrovec, J. (2017). Contrasting effects of ploidy level on seed production in a diploid–tetraploid system. AoB Plants, 9(1), plw077. https://doi.org/10.1093/aobpla/plw077
Murtadha, H. M., Maranville, J. W., Clark, R. B., & Clegg, M. D. (1989). Effects of temperature and relative humidity on growth and calcium uptake, translocation, and accumulation in sorghum. Journal of Plant Nutrition, 12(5), 535–545. https://doi.org/10.1080/01904168909363972
Norden, E. H., Lyrene, P. M., & Chaparro, J. X. (2020). Ploidy, fertility, and phenotypes of F1 hybrids between tetraploid highbush blueberry cultivars and diploid Vaccinium elliottii. HortScience, 55(3), 281–286. https://doi.org/10.21273/HORTSCI14597-19
Ochmian, I. D. (2012). The impact of foliar application of calcium fertilizers on the quality of highbush blueberry fruits belonging to the ‘Duke’ cultivar. Notulae Botanicae Horti Agrobotanici Cluj-Napoca, 40(2), 163–169. https://doi.org/10.15835/nbha4028058
Olmedo, P., Zepeda, B., Rojas, B., Silva-Sanzana, C., Delgado-Rioseco, J., Fernández, K., Balic, I., Arriagada, C., Moreno, A. A., Defilippi, B. G., & Campos-Vargas, R. (2021). Cell wall calcium and hemicellulose have a role in the fruit firmness during storage of blueberry (Vaccinium spp.). Plants, 10(3), 553. https://doi.org/10.3390/plants10030553
Park, S. W., An, S., & Kwack, Y. (2020). Changes in transpiration rates and growth of cucumber and tomato scions and rootstocks grown under different light intensity conditions in a closed transplant production system. Journal of Bio-Environment Control, 29(4), 399–405. https://doi.org/10.12791/KSBEC.2020.29.4.399
Rojas‐Barros, P., Wernow, J., Workmaster, B. A., Zalapa, J., Devi, J. M., & Atucha, A. (2025). Fruit cuticle thickness and anatomical changes in pedicel xylem vessels influence fruit transpiration and calcium accumulation in cranberry fruit. Physiologia Plantarum, 177(1), e70036. https://doi.org/10.1111/ppl.70036
Sharma, P., Janaagal, M., Sheoran, A. R., Luhach, A., Sharma, D., & Sharma, C. (2024). Multifaceted responses to heat stress in plants: A review of the morpho-physiological, biochemical and anatomical changes. Asian Research Journal of Agriculture, 17(4), 1059–1071. https://doi.org/10.9734/arja/2024/v17i4620
Spiers, J. M. (1983). Elemental leaf content and deficiency symptoms in rabbiteye blueberries: 2. Calcium and magnesium. Journal of Plant Nutrition, 6(12), 1073–1086. https://doi.org/10.1080/01904168309363169
Strik, B. C., & Vance, A. J. (2015). Seasonal variation in leaf nutrient concentration of northern highbush blueberry cultivars grown in conventional and organic production systems. HortScience, 50(10), 1453–1466. https://doi.org/10.21273/HORTSCI.50.10.1453
Strik, B. C., & Vance, A. J. (2019). Highbush blueberry cultivars differ in the relationship between seed number and berry weight during the harvest season. HortScience, 54(10), 1728–1736. https://doi.org/10.21273/HORTSCI14198-19
Stückrath, R., Quevedo, R., de la Fuente, L., Hernández, A., & Sepúlveda, V. (2008a). Effect of calcium foliar application on the characteristics of blueberry fruit during storage. Journal of Plant Nutrition, 31(5), 849–866. https://doi.org/10.1080/01904160802043197
Stückrath, R., Quevedo, R., de la Fuente, L., Hernández, A., & Sepúlveda, V. (2008b). Effect of foliar application of calcium on the quality of blueberry fruits. Journal of Plant Nutrition, 31(7), 1299–1312. https://doi.org/10.1080/01904160802135076
Taber, S. K., & Olmstead, J. W. (2016). Impact of cross- and self-pollination on fruit set, fruit size, seed number, and harvest timing among 13 southern highbush blueberry cultivars. HortTechnology, 26(2), 213–219. https://doi.org/10.21273/HORTTECH.26.2.213
Tadesse, T., Nichols, M. A., Hewett, E. W., & Fisher, K. J. (2001). Relative humidity around the fruit influences the mineral composition and incidence of blossom-end rot in sweet pepper fruit. The Journal of Horticultural Science and Biotechnology, 76(1), 9–16. https://doi.org/10.1080/14620316.2001.11511319
Tamada, T. (1998). Nutrient deficiencies of rabbiteye and highbush blueberries. Acta Hortic., 241, 132–138. https://doi.org/10.17660/ActaHortic.1989.241.20
Tanguilig, V. C., Yambao, E. B., O’Toole, J. C., & De Datta, S. K. (1987). Water stress effects on leaf elongation, leaf water potential, transpiration, and nutrient uptake of rice, maize, and soybean. Plant and Soil, 103, 155–168. https://doi.org/10.1007/BF02370385
Tromp, J., & Oele, J. (1972). Shoot growth and mineral composition of leaves and fruits of apple as affected by relative air humidity. Physiologia Plantarum, 27(2), 253–258. https://doi.org/10.1111/j.1399-3054.1972.tb03610.x
Vance, A. J., Jones, P., & Strik, B. C. (2017). Foliar calcium applications do not improve quality or shelf life of strawberry, raspberry, blackberry, or blueberry fruit. HortScience, 52(3), 382–387. https://doi.org/10.21273/HORTSCI11612-16
White, P. J. (2001). The pathways of calcium movement to the xylem. Journal of Experimental Botany, 52(358), 891–899. https://doi.org/10.1093/jexbot/52.358.891
White, P. J., & Broadley, M. R. (2003). Calcium in plants. Annals of Botany, 92(4), 487–511. https://doi.org/10.1093/aob/mcg164
Wójcik, P. (2005). Response of ‘Bluecrop’ highbush blueberry to boron fertilization. Journal of Plant Nutrition, 28(11), 1897–1906. https://doi.org/10.1080/01904160500306425
Yang, F. H., Bryla, D. R., & Strik, B. C. (2019). Critical temperatures and heating times for fruit damage in northern highbush blueberry. HortScience, 54(12), 2231–2239. https://doi.org/10.21273/HORTSCI14427-19
Yang, F. H., DeVetter, L. W., Strik, B. C., & Bryla, D. R. (2020). Stomatal functioning and its influence on fruit calcium accumulation in northern highbush blueberry. HortScience, 55(1), 96–102. https://doi.org/10.21273/HORTSCI14482-19






